
Winter Juvenile Tree Dieback (WJTD) – a Problem in Pistachio in the San Joaquin Valley
By Craig Kallsen, Citrus and Pistachio and Blake Sanden, Soils and Water: UCCE Farm Advisors Kern County
More WJTD this spring (2019) in the southern San Joaquin Valley
Winter juvenile tree dieback (WJTD) was a common problem this spring in a number of areas in the southern San Joaquin Valley (SJV) and points north. Symptomatic trees ranges from a fraction of a percent to about 50% of the trees in parts of some blocks. Symptomatic trees included those on PG1, UCB1 seedlings and clones. Affected scions on these rootstocks included Kerman, Golden Hills, Peters and Randy. In all blocks were temperatures were available, the primary cause appears to be a cold snap during early to mid-November. Air temperatures in this calendar period ranged from daytime highs in the high-to-mid-70s °F with nighttime low air temperatures as low as 26 °F.
This newsletter is a compilation of a decade’s worth of observations related to this problem. Much of the information presented has not been developed from rigorous replicated and randomized scientifically designed trials. There remains much to prove and much to learn. Some suggestions for reducing the occurrence and severity of WJTD are discussed.
General Description/Symptomology of the Problem
Juvenile pistachio trees appear to be in excellent health, vigorous and healthy, as they go into fall dormancy. Winter juvenile tree dieback typically occurs in the time-period between late October and mid-December in trees from one to seven years old, although trees third to fifth leaf are most at risk. The major cause of WJTD is attributable to freeze damage, although this has not been proven and the extreme variability in symptoms within an orchard and neighboring orchards remains puzzling. A tree with WJTD is often immediately adjacent to an unaffected tree, although typically, affected trees appear in clumps. Trees uninjured or injured the previous year may be either injured or not injured the subsequent year. Orchards that would appear at extreme risk show no WJTD, while neighboring orchards, with lower apparent risk, may be severely affected. As is discussed below, there are contributing factors, which makes trees more susceptible to WJTD.
Reported incidences of WJTD increase greatly soon after extended freeze events in October, November and into the third week in December, especially so if the freeze occurs suddenly before the trees are hardened off. Some symptoms vary with the degree of cold experienced and when during the fall the cold temperatures occur. Kerman, Peters, Randy, Kalehghouchi and Golden Hills cultivars growing on UCB1 seedlings, UCB1-clonal material, and PGI rootstocks have been affected.
Winter juvenile tree dieback (WJTD) is not a new phenomenon. There is a picture that Joseph Maranto, my predecessor as pistachio farm advisor in Kern County, published in the early 1980s of a tree he suspected of being damaged by cold temperatures. To confound the problem, WJTD has occurred, rarely, when below freezing temperatures were not present and this observation is discussed in more detail below. Juvenile Pistachio Freeze Damage can be lumped into two broad categories:
Mild
Severe
Symptoms of WJTD
1. Mild WJTD – Early spring leaf-out
First through eighth leaf pistachio trees, and perhaps older, having rootstocks with P. integerrima heritage, such as UCB1 and PG1, can show freeze damage in late October or early November if growing vigorously, even in geographical areas where soil salinity is not a problem. The current hypothesis is that temperatures sufficient to cause mild symptoms on vigorous trees may be in the range of 27 to 31°F. Even in the coldest areas of Kern County, temperatures lower than this are rare in late October through mid-November.
Reported incidences of mild freeze damage symptoms increase greatly soon after mild freeze events in late October and November before trees show much dormancy. Typical symptoms include black mold, bark wetness and white gum on the lower scaffold branches and trunk above the graft union. These symptoms, especially, are prevalent if the trees have not yet developed any degree of dormancy. If the freeze occurs before leaf drop, which is usually the case with mild WJTD, the leaves of frozen branches remain on the tree and are very distinctive after the leaves of unfrozen branches lose their leaves. Usually pruning crews are the first to find this type of injury (Photo 1).

Photo 1. Pistachio tree showing typical symptoms of attached leaves in Winter Juvenile Tree Dieback (WJTD). Leaves remain attached if the frost event occurred before leaf drop (typically in late October or early November).
Trees showing these symptoms in the fall, often leaf out and flower (if old enough), earlier than unaffected trees in the spring. Leaves near the end of branches may be twisted and malformed. With the first string of very hot days in May, affected outer branches may die back, unable to keep up with the transpiration requirements due to damage to the vascular tissue. However, frequently, little or no die back occurs and the tree will grow normally. Winter Juvenile tree dieback should not be confused with the “normal” characteristic of the male cultivar Randy, which tends to hang onto its leaves longer in the fall than does Golden Hills.
2. Severe WJTD – late spring leaf-out.
Severe WJTD appears to occur after early November through late December. Usually temperatures do not drop low enough, even at low elevations, prior to mid-November to cause severe WJTD. As the trees gain greater dormancy from October through early December, colder nighttime temperatures are required to cause WJTD, but rootstock, soil salinity/permeability, and other crop-related factors, appear to influence just how cold temperatures must be to cause the problem. Approximate observed threshold air temperatures cold enough to cause WJTD through fall in some orchards are displayed in Figure 1. Air temperatures from an actual orchard where WJTD was severe are shown in Figure 2.

Figure 1. Low temperatures (measured at a height of approximately 4 ft. above the ground) on various dates where severe WJTD occurred to moderately hardened-off juvenile pistachio trees in various low-elevation, salt-affected orchards in the southern San Joaquin Valley. This figure should be considered very preliminary and data are taken from freeze events over the past ten years.

Figure 2. The chart above was prepared from temperature data collected in two 5th-leaf orchards during the night of December 6-7, 2012. On several other nights in early December, temperatures fell to similar levels. The trees in the warm orchard showed no WJTD, while those in the cold orchard had approximately 20% severe WJTD.
Trees severely affected later by WJTD, in mid-November through mid-December, normally do not exhibit attached leaves, or as much mold growth and light gumming after the freeze event. These trees, however, are more heavily damaged than those showing only mild symptoms in the early fall. Typically, symptoms are not apparent until spring. Affected first and second leaf trees may simply die without pushing any new growth. In trees third-leaf or older, the new growth, typically may push only from the lower scion or rootstock and this growth occurs later than in unaffected trees. Clear gum, may be present on the scion, but normally exudes from the lower rootstock and graft union. The bark may turn black with time, but will appear dry. The dead tissue will rapidly become colonized by secondary fungal pathogens such as species from Phomopsis, Botryosphaeria, Fusarium and Alternaria.
Some affected trees show minimal dieback, while many trees in the worst affected blocks die back to the rootstock, or the entire tree is killed. When air temperatures are very cold, below 25 degrees F, bark and cambial tissue may be killed forming sunken cankers, especially on the south and west sides of the tree. We suspect the reason that there is more tissue damage on the south and west sides of the trunk is that this is the more sun-exposed part of the trunk. After a cold night, the sun-exposed sides of the trees warms rapidly and the damaged cells thaw rapidly, resulting in more cellular damage. As is the case with the mild symptoms of WJTD, Kerman, Peters, Kalehghouchi, Golden Hills and Randy cultivars growing on UCB1 seedlings, UCB1-clonal material, and PGI rootstocks have been affected . Peters appears more susceptible to damage than Kerman. In one orchard near Delano, less than two percent of the damaged trees were Kerman the rest were Peters.
Severely damaged trees may only leaf-out from the lower scion and/or rootstock of the trunk in May, well after the unaffected branches on the same tree of different trees have leafed out. Observations suggest that for trees to be severely damaged by frost after late November, temperatures usually fall below 25 ºF, with temperatures below freezing for an extended part of a night or multiple nights. Trees appear to be dormant enough by late December that they are no longer easily damaged by freeze events.






Additional Observations related to WJTD are as follows (and are addressed individually in the text below):
(A) Rootstocks with 100% P. integerrima heritage (such as PG1) and clones developed from UCB1 selections (referred to as ‘UCB1 clones’) have a higher percentage of damage than occurs with trees produced from seeds from the UCB1 cross (a selected female P. atlantica by a selected male P. integerrima) referred to as ‘UCB seedling’ trees.
(B) Topographically low-elevation orchards have had most of the WJTD, but damaged trees, often, are not in the lowest areas of a given orchard.
(C) Soil chemical and physical characteristics, such as high sodium, EC, silt, and presence of hardpans and perched water tables increase WJTD.
(D) The scion is usually more adversely affected than the rootstock, but the entire tree may be killed.
(E) A tree with WJTD is often immediately adjacent to an unaffected tree, although typically, affected trees appear in clumps.
(F) Various degrees of dark streaking may be found in the trunks (usually the scions) that appear to be similar to those caused by Verticillium wilt, although as of this writing Verticillium isolations have not been completed.
(A) Rootstocks with 100% P. integerrima heritage (such as PG1) and UCB1 clones have a higher percentage of damage than occurs with UCB1 seedling trees.
The severity of the WJTD problem is associated with the rootstocks currently used in the pistachio industry. Specifically, WJTD is related to the amount of P. integerrima in a rootstock’s heritage. Rootstocks planted since the early 1980s are genetically either 100 percent P. integerrima, such as PG1®, or 50 percent P. integerrima, such as UCB1. Trees grafted to rootstocks with 100 percent P. integerrima heritage, have shown poorer frost tolerance than UCB1 seedlings in a number of orchards grafted to both types of rootstocks and in randomized and replicated UC rootstock trials. This observation does not mean that we should not use rootstocks with P. integerrima heritage. The discovery by University of California researchers that Pistacia integerrima was resistant to Verticillium Wilt disease was of tremendous importance to the California pistachio industry. Rootstocks with P. integerrima have been and will continue to be a critical component of our industry here and have demonstrated the ability to produce very high yields. However, P. integerrima is not a perfect match for the San Joaquin Valley, which is not surprising, since this species of pistachio is adapted to a subtropical area of the Indian subcontinent. Winter juvenile tree damage, also, is more severe and affecting a higher number of trees in affected orchards in UCB1 clonal orchards than UCB1 seedling orchards, probably due to the greater vigor, on average, of the clones. Pistacia vera scions (i.e. the commercial nut bearing portion of the tree) such as ‘Kerman’ grafted onto rootstocks having P. integerrima heritage or onto vigorous UCB1 clonal rootstocks, do not appear to be as well adapted to pick up environmental cues that winter is approaching as they would if growing on their own roots and not grafted to these rootstocks. These cues include shorter day length and reduced nighttime temperatures.

Note picture of a pistachio tree having a P. integerrima rootstock taken Dec. 10, 2010. The tree was well watered until this date. Note that the scion leaves have dried indicating some degree of dormancy in the scion, whereas the rootstock sucker leaves remain green and succulent. What is happening at the graft union as the P. vera scion goes dormant and the P. integerrima rootstock does not ?
If water is available, PG1 and UCB1 clones continue to grow vigorously into the fall and winter, since temperatures in the SJV, normally, remain warm throughout the fall. In fact, trees on all of our commonly used rootstocks, no matter which one, do not appear to go fully dormant until the end of December. Evaluations in randomized and replicated UC rootstock trials have shown that UCB1 seedlings have greater salt tolerance than PG1, which may be an important factor in the reduced incidence of severe WJTD symptoms in UCB1 seedling orchards. The effect of salinity on WJTD is discussed further in Section C below.
(B) Topographically low-elevation orchards have had most of the WJTD, but, often, damaged trees are not in the lowest areas of a given orchard.
Winter juvenile tree dieback is not new to the San Joaquin Valley. In the past, we have called this problem freeze damage. What is new is the extent of the problem. One explanation for the increase in WJTD hypothesis that thousands of new acres of pistachio trees were planted over the recent past in low-elevation areas of the San Joaquin Valley at the same time that we experience more low-rainfall years. Cold air drains into low elevation areas making them colder than higher ground and low rainfall translates into reduced fog. The absence of fog means drier air with low dew point temperatures. Dry air means air temperatures get colder faster in the evening once the sun goes down, resulting in lower lows and longer durations of cold temperatures, with much more potential to damage vigorously growing young trees. The most obvious explanation for WJTD during the winter is freeze damage. Reports of WJTD always spike shortly after or in the following spring after a major early-fall freeze event. Investigations of WJTD-affected trees for disease (with the possible exception of Verticillium wilt –see Section E) and the soil for harmful levels of heavy metals, have found nothing suggesting these are causal factors. Juvenile trees die at both low elevations and high elevations, as long as freezing temperatures are present.
A further problem with growing pistachios in these low-elevation, former lake beds is that salts, such as formed from sodium, boron and chloride, also tend to collect in low-lying areas because ofirrigation drainage and evaporation. The association of WJTD with deflocculated, silty, saline soils with possible hardpans is discussed in Section C below.
(C) Soil chemical and physical characteristics, such as high sodium, EC, silt, and presence of hardpans and perched water tables appear to increase WJTD.
The chemistry of the soil water and the amount of water in the root zone appear to be involved in the severity of WJTD but separating the effects of salinity from late-season soil-water storage is difficult. Soils do not have to be saline for WJTD to occur; however, as mentioned above, damage appears to be more common and more severe in salt-affected orchards. Areas with high salinity are usually associated with silty, poorly draining soils and shallow hardpan or high water tables. The element making up the salt, also, appears to be important. Low calcium in plants has been associated in the literature with increased sensitivity to frost for a number of plant species including trees, and, often in the SJV, low calcium means high sodium. However, plants do not appear to be poisoned by sodium. Differences in calcium and sodium concentrations in trunk tissue between affected and unaffected pistachio plant tissues has been found in our testing, but this appears to be an artifact of freeze damage. Since pistachio excludes salts from its leaf cells and appears to be able to mobilize absorbed sodium from the plant back to the soil, we have been unable to establish much difference in plant sodium concentration between plant parts of affected and unaffected trees shortly after a freeze event in the fall when the trees begin showing the first symptoms. However, the concentration of sodium is much higher in the bark and outer wood near the cambium of the trunk of severely injured WJTD trees in May after leaf-out. This observation suggests that sodium is leaking out of damaged xylem vessels in the trunk and into the wood and bark as the flow in the xylem responds to the increased transpirational demand on the tree in the spring, and is not higher in these tissues prior to the freeze event.
Winter juvenile tree dieback has been associated with areas of an orchard altered by recent or historical human activities. Areas of orchards where roads or feedlots had existed prior to the establishment of the orchard, or where pipelines were dug across the orchard, or significant grading had occurred, all appear to increase the probability of trees damaged by WJTD. In the photos below, the difference in weed growth marks the transition in field from where a significant depth of topsoil was graded from the orchard to a depth of about 15 inches. The area with weeds is where the original topsoil remains. The area with the weeds showed no damage from WJTD, while the area not having weeds, had WJTD symptoms, many severe, in about 15% or more of the trees over multiple years.

This photo shows an area in the graded and weedless area of the orchard. The smaller trees in the bottom photo were replants or grafted suckers from similarly affected trees by WJTD the previous year.

This photo shows the abrupt change in weed growth within the orchard because of land grading conducted before the orchard was planted.
The degree of susceptibility of juvenile trees to frost may be more of a function of increased wetness in the root zone as opposed to direct toxicity of sodium or other salts. Trees that are drought stressed in fall, prior to frost event, are likely more resistant to frost damage (Sperling et al., 2016). The existence of layers in the soil that are more impermeable to water, or soils with textures that are high in silt, appear to increase the hazard of WJTD, especially in soils were the concentration of salt in the soil and irrigation water is already high. These kinds of soils are difficult to drain and leach, and salts of several elements accumulate. If sodium is the major cation present, the soils will lose structure making the problem worse. Soil water that has high EC, that is higher salinity, will have a greater osmotic pressure (or lower osmotic water potential) meaning that plants will have greater difficulty absorbing and transpiring water. These soils never truly dry out. Thus roots in saline soil will tend to be wetter and more likely to suffer from lack of oxygen (that it, hypoxia) in the fall as they begin to go into the winter rest period, even if impermeable hardpans do not exists. Trees never go into full dormancy and thus still metabolize during the winter. To metabolize, plants require oxygen to be present in the soil. If that oxygen is not present, it may result in a tree much more susceptible to WJTD.
Elevated sodicity, decreased available calcium, and high water holding capacity, are characteristics very consistent with a large number of orchards and of small alkali areas (that is “hotspots”) within larger orchards across the San Joaquin Valley. A detailed examination of one such “hotspot” within an orchard already somewhat yield compromised by high salinity across the entire orchard, was conducted by Irrigation and Soil Farm Advisor Blake Sanden (UCCE, Kern County) beginning in 2011.
Investigation of a WJTD “hotspot”
Soil samples and measurements of soil magnetic conductance were taken along a transect of 15 trees from the center of the affected area with considerable tree death and decline through trees with intermediate damage outward to a vigorous unaffected area of the orchard. Figure 3 demonstrates how the sodium adsorption ratio (SAR) decreased with increasing distance from the center of the hotspot. The SAR is a measure of the amount of sodium compared to calcium in the soil. In general, calcium tends to improve soils structure and sodium to diminish it.

Figure 3. The sodium absorption ratio at three locations in relation to the irrigation hose ratio at different soil depths (upper left, upper tight and lower left charts) and as an average (lower right chart) with increasing distance from the worst affected trees in the center of the hotspot (marked as 0 on the x-axis).
Soil magnetic conductance readings were made using a Geonics EM38-DD along the same transect where the SAR values in Figure 3 were measured. The EM38-DD is a device about 40 inches long. It sends an electromagnetic pulse from one end of the instrument to the other – measuring the change in the signal through a quarter-sphere of the root zone soil about 40” wide and 40” deep. This
12
“conductance” reading is often called an apparent soil electroconductivity (ECa) which is usually correlated to the lab soil sample salinity, EC. Nevertheless, this conductance is affected by water content, soil mineralogy and density as well as soil salinity. EM38-DD readings consist of a magnetic pulse in the horizontal (EMh) and vertical (EMv) planes basically integrating a “bulk conductance” for a soil volume representing a quarter sphere in volume with a diameter of 1 to 1.2 meters depending on soil texture and water content.
The “usual” assumption in using the EM38-DD, is that the water content will be much more uniform at any given measurement period than the differences in soil salinity. Therefore, the variation in the readings are attributable, mostly, to salinity differences. However, under the conditions of this study, water content was not uniform across the area of measurement. Soil water content tended to be higher where salinity and poor drainage was present. In this instance and as used in these hotspots, a given soil conductivity measurement provided by the EM38-DD was considered an “integrated’ measurement of both salinity and the amount of water held in the soil. Root hypoxia is a condition of plant roots where oxygen concentration is insufficient for normal metabolism, while anoxia occurs when no oxygen is available. A higher soil conductance measurement then, is an indication of both higher soil moisture and higher salinity, both of which contribute to higher plant salt stress and root hypoxia. The EM38-DD measurement displayed in Figure 3 showed a similar pattern to the SAR measurements in Figure 2. Soil conductance readings, a measure of EC and possible root hypoxia/anoxia, were higher the closer the instrument was to the center of the “hotspot” (see the top left and top right charts in Figure 4). When the EM38-DD raw readings (upper left and right charts in Figure 4) are compared to the EC measurements resulting from separate soil samples taken along the same tree transect (see lower right chart in Figure 4), the same pattern is apparent with decreasing EC values with increasing distance from the hotspot.
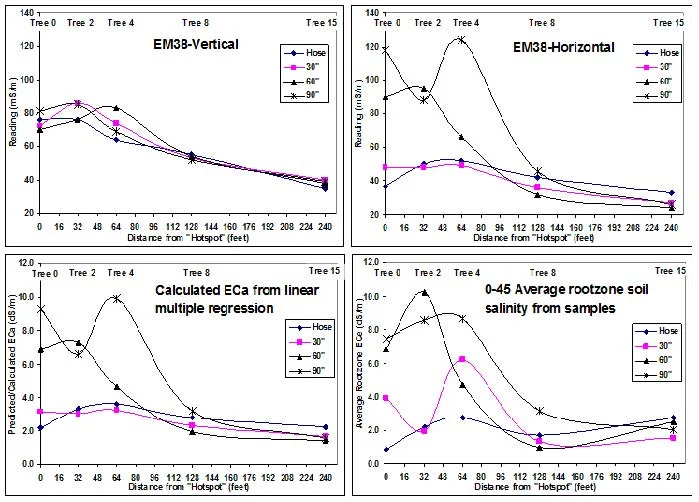
Figure 4. EM38-DD readings (top left and right charts) and saturation extract laboratory electrical conductivity (EC) (bottom right chart) with increasing distance from the worst affected trees in the center of the hotspot.
These data, from this hotspot and similar hotspots examined in 2011, show that soil physical and chemical characteristics affected the severity of WJTD. In this particular study, involving several hotspots, WJTD susceptibility ceased to be a problem when average root zone soil salinity was
ECe < 6 dS/m, soluble Na < 40 meq/l (920 ppm) and Na/Ca ratios < 15 at a distance of 60 to 90 inches from the drip hose. This roughly corresponds to an EM38-DD vertical and horizontal summed reading of about 80 milli-Seimens/meter (mS/m). These values, while informative, should not be considered “thresholds” transferable to other orchards and other freeze events. Winter juvenile tree dieback has occurred under a wide range of soil types, soil chemical and physical characteristics, orchard topography, air temperature minimums, fall calendar timings, and rootstock and scion selections. However, elevated soil EM38 readings, sodium to calcium ratios, and salinity should be considered as increased risk factors for future development of WJTD during freeze events.
The use of the EM38 in a similar way, but for characterizing salinity and its effect on nut yield, is another area of active investigation. In a separate 3 year regional study (2014-16) of 10 salt-affected orchards in Kern County (using 3 to 4 sites in each orchard from “good” to “bad”) Sanden found a better correlation of pistachio yield decline with EM38 bulk conductance than using the traditional soil salinity lab ECe. The yield decline threshold started at a similar summed bulk conductance of 80-120 mS/m as shown in Figure 4.
To complicate an understanding of WJTD further, symptoms of this dieback were found in some years in orchards where no apparent freeze event occurred. Winter juvenile tree dieback, without an apparent freeze event are rare, with only three instances noted over the past decade. In two of the cases, the soils in these orchards were moderately saline, heavily manured and likely subjected to intervals of waterlogging in a wet winter. What caused the WJTD in these frostless orchards remains unexplained. However, there is a limit to how much manure can be disposed of around a growing crop such as
14
pistachio and that limit is likely is dependent on soil texture, salinity, drainage, degree of fall/winter water-logging and other factors, most or all of which are not known. The decomposition of manure requires oxygen, and produces ammonia and other gases, which in high concentration could result in root hypoxia/anoxia. Perhaps, hypoxia could be severe enough to cause WJTD in saturated, deflocculated soils, without an associated freeze event.
(D) The scion (that is the nut-producing top of the tree) is usually more adversely affected than the rootstock; but the entire tree may be killed;
Some qualitative starch analysis of the rootstock of unaffected and trees with WJTD, conducted after leaf-out in 2011, demonstrated that unaffected trees contained almost no starch below the graft union either in the trunk or roots, while trees severely impacted by WJTD had high concentrations of starch in the rootstock, both above and in the large roots below ground. This suggests that all of the trees in the orchard were healthy going into the winter, and had stored large amounts of starch in the scion trunk, roots and above ground portions of the rootstock. The trees not damaged by frost were able to mobilize these starch reserves in the spring and use them to produce a new leaf canopy. Starch stored in trees that had died back, and now had too few unfrozen growing points left, remained stored in the rootstock instead of used to make a new leaf canopy. In some frost-damaged trees, there was a clear line of demarcation between no starch in the scion directly above the graft union and a large concentration of starch in the rootstock directly below the graft union. These trees appeared to have lost the ability to translocate starch from the rootstocks to the scion.

Plant tissues treated with Lugol’s solution will turn a dark color if starch is present. Note that starch appears to be completely absent in this unaffected tree treated with Lugol’s solution, indicating that the tree, apparently, was able to mobilize starch reserves in the rootstock for new canopy growth in May.

Note the well-marked line of demarcation (dotted black line) in the wood under the bark at the graft union in this WJTD-affected tree in May. This line, also, marks the graft union. The graft union of this tree looked identical to that of the tree in the picture on the right until stained with Lugol’s solution. The dark stain below the graft union indicates presence of starch, as stained with Lugol’s solution below the graft union, with little starch remaining above the union in the scion.
The clear line of demarcation at the graft union also suggests that the graft union, the place where P. integerrima melds with P. vera, is a weak point with respect to frost tolerance. In Montana and Minnesota, where the temperatures get very cold, it is very difficult for any grafted landscape plants to survive the winters. In cold climates, the graft union is a point of weakness in susceptibility to frost. Similar examples exist in backyard lemon trees in Bakersfield. It is common for the lemon scion to have been killed in a frost event, and all that remains to the homeowner is a vigorous, thorny rootstock tree. The xylem (i.e. the water conducting tissues) are the most susceptible structures in the trunk to frost damage and the xylem vessels most at risk in the tree appear to be those at the graft union. In freeze-damaged orchards, rootstocks regrow vigorously with the remnants of the scion or minus the scion, which has died. Kallsen has been involved in a number of seedling breeding plots, where P. vera are grown on their own roots. Some of these seedling selection trials have been located in low-elevation cold areas. No P. vera tree on its own roots has exhibited WJTD in these trials.
If freeze damage occurs largely at the graft union, tree wraps should mitigate damage, although to our knowledge, they are not being used for this purpose (see info on tree wraps in later section below).

Note gumming from graft union.

Note UCB-1 rootstocks growing absent the scion. When the scion dies as a result of WJTD, the rootstock often regrows vigorously, suggesting the grafted tree is more frost sensitive than the rootstock alone.
(E) A tree with WJTD is often immediately adjacent to an unaffected tree, although typically, affected trees appear in clumps.
Even trees on clonally produced rootstocks show marked differences in WJTD within an orchard. This observation suggests that the different genetics in seedling rootstock are not the primary reason for the variability in tree damage in an orchard. More likely explanations relate to the microclimatic air-temperature variability within the orchard during the actual freeze event, the past growth performance of individual trees within the orchard, and as described above, to variations in soil physical and chemical characteristics.
Commonly, fall freeze events begin with a large mass of cold air moving into the SJV from a front originating in Alaska. However, these autumn fronts, normally, do not bring in air temperatures sufficiently cold to cause freeze damage directly. Freeze damage normally occurs in response to what happens after the arrival of the colder air mass and once the wind and clouds dissipate. On clear and windless nights following the arrival of the cold front, radiative cooling from the ground to air begins. As the air near the ground continues to lose heat, it can become several degrees colder than the air above it and this transfer of energy is not uniform. Warmer air near the ground will push skyward in bubbles. Thus, on nights like these there is little uniform air mixing and the orchard atmosphere is composed of ‘bubbles’ of air of different temperatures. Air temperatures around trees may vary considerably, depending on the microclimate (i.e. air bubbles) that are in contact with a given tree throughout the freeze event. Typically, in late October or early November, the difference between damage and no damage can be as little as from 32 degrees F (no damage) to 31 or 30 degrees (extensive damage). All of the trees in an orchard would freeze if average temperatures dropped sufficiently, but air temperatures like these, fortunately, do not occur in October and November in the SJV, and, at worst, are borderline for producing tree damage.
When looking at an orchard, all we can see are the parts of the trees above ground. Because of tree training and pruning, most of the aboveground portions of trees have a very uniform appearance. However, the root mass and root proliferation of each tree, even those on clonal rootstocks, may vary considerably. A tree that take a bud early and grows vigorously may have a much more extensive root system than its neighbor has and have a variable propensity for freeze damage due to differences in tree vigor, soil water availability, soil salinity and root hypoxia.
(F) Various degrees of dark streaking may be found in the trunks (usually the scions) of affected trees, which appear to be similar to those caused by Verticillium wilt, although as of this writing Verticillium isolations are not yet completed.
The association of Verticillium and WJTD is an active area of investigation. Currently, Drs. Themis Michailides and Florent Trouillas, UC plant pathologist, are investigating the streaks to determine if they are caused by Verticillium wilt or, perhaps, freeze damage. Verticillium infections are not unusual in trees in the SJV, but the natural levels of tolerance/resistance in our UCB1, PG-1 and Platinum rootstock generally prevent it from causing economic harm. If the streaking is found to be caused by Verticillium wilt, Dr. Michailides has hypothesized trees with more Verticillium infections may be predisposed to freeze damage or, possibly, that freeze-damaged trees are more susceptible to Verticillium infection.
Why trees dieback
Generally, the likely cause of the dieback is damage to the water-conducting vessels initiated by cold temperatures. The tree dies back due to dehydration, or inability to supply water for maintenance and growth of the tree.
Typically, when a plant part is damaged by something like frost, mechanical damage, an herbivore or insect infestation; ethylene (a plant growth regulator) is released by injured plant cells. Ethylene induces a stress response in the plant, which, in addition to affecting many other physiological/metabolic processes, results in cells adjacent to the wound site sealing off the damaged area. For example, parenchyma cells associated with the xylem create tyloses, which affectively seal off the xylem, preventing excessive loss of water by the plant.
Cells of trees foliarly treated with ethephon release ethylene as this chemical penetrates the plant after spraying. Interestingly, applying ethephon, in a commercial product, to the canopy and trunk of a pistachio tree at concentrations greater than 500-ppm active ingredient in November before it is dormant, appears to mimic the symptoms of gumming and early leaf out, seen in frozen trees. If a tree is frozen, but not so badly that the upper canopy is killed, we would expect to see symptoms, such as a stress-related early leaf out, since ethylene gas being released by damage cells affect living buds. Trees that do not show the early, mild symptoms but only sprout from the lower trunk in May, may have been damaged so badly by frost, that the ethylene response was overwhelming, resulting in the death of the scion and upper rootstock. When ethephon was sprayed, experimentally, on trees in late January through mid-February, after maximum dormancy was achieved, the trees demonstrated early leaf out comparable to that when treated with oil in the same time frame, but no WJTD symptoms. Please note that ethephon is not labeled for use in pistachio, for any purpose.

Damaged shoot with dead vegetative and flower buds in spring.

Earlier leaf out than neighboring unsprayed trees.

Trunk showing gumming.
Trees that are less than one-year old and up to second leaf may be killed outright by freeze events. The rootstock dies first, followed shortly by the scion. The trees usually begin dying back in May with the first warm temperatures and the dry or drying leaves will be obvious from a distance. At this time, if you scratch the bark, the bark of the rootstock will be gray to black, while that of the scion will still be green (that is still alive). There appears to be insufficient root mass and carbohydrate storage in first-year trees to support regrowth of suckers, and the rootstock dies. Older trees, with their more extensive root systems, appear to have sufficient carbohydrate reserves to survive and push new suckers, which the grower can graft to, to regrow the tree. In general, it may be easier to encourage dormancy in very young trees, since they have not yet developed extensive root systems, and are more susceptible to drought-induced reductions in vigor and earlier dormancy.
As mentioned above, excessive tree vigor going into fall appears to have a strong association with WJTD. Trees older than seven or eight years appear to be largely immune from the problem, but, of course, if temperatures get cold enough, anything can freeze. Perhaps bearing large yields of nuts or prolific amounts of pollen reduces tree vigor, or something about the rhythmicity of the bearing cycle once fully established, allows trees to become dormant earlier. As trees begin bearing, the bark thickens, and this thick bark may resist frost damage better. The Kalehghouchi cultivar, probably, is the most vigorously growing cultivar grown. Kaleghouchi, at a couple of sites, where it was growing adjacent to Kerman and Golden Hills blocks, appeared to be more susceptible to WJTD.
In general, observations suggest that trees fully irrigated late into the season appear to be more at risk of WJTD. Wetter soils appears to be associated with increased severity of WJTD, but why this occurs is a subject of debate. Areas of the orchard with poor infiltration or shallow hard pans can hold water later into the season, perhaps allowing more vigorous freeze-susceptible growth compared to trees “hardened” by drought. Drought hardening has been shown to decrease the temperature threshold at which rootstock seedlings freeze. However, wet, poorly structured soils may lead to root hypoxia in the fall, when evapotranspiration rates plummet but the trees are not yet dormant. In 2019, some trees growing immediately adjacent to a large canal were severely damaged by an early frost in November, probably related to measured water seepage into the field at a depth of three to four feet in the soil profile. Again, whether this water promoted high vigor going into the fall, or was associated with root hypoxia/anoxia as the trees went into winter dormancy is unknown. Juvenile pistachio trees show very little sign of slowing down on their own if sufficient soil-water is available. In the fall in the SJV, there is often very little transition to cooler winter temperatures. Daytime high air temperatures can easily be in excess of 80º F in October and early November and suddenly crash to below freezing at night. The variability of the degree of WJTD in an orchard may reflect differences in individual tree vigor within the orchard.
What the Grower Might Be Able to Do to Reduce the Incidence of WJTD
Choice of rootstock
UCB1 seedling rootstock (that are by heritage 50% P. integerrima and 50% P. atlantica ) suffer less WJTD than either PG1 (100% P. integerrima heritage) or UCB1 clonal rootstock. UCB1 has been shown to be more frost tolerant than P. integerrima. The reason that UCB-1 clones have demonstrated more instances of severe WJTD that the seedlings, is thought to be that the clones were chosen for their vigor, which appears to much higher than the average UCB1 seedling However, choice of rootstock is not enough. Records show, that of all of the visits made to WJTD-orchards, no UCB1 seedling orchards suffered more than 1% damage, and most were much less than this. Damage to some PG1 and UCB1 clonal orchards, was as high as 25% or more. However, all of our commercial rootstocks are vigorous and additional measures will be required to prevent or reduce WJTD.
PG1 is a seedling rootstock as, obviously, are UCB1 seedlings. Each PG1 or UCB1 rootstock is, to some degree, genetically unique, unlike a clonal rootstock. Genetic differences between individual trees may account for some of the variability seen in WJTD-damaged orchards. Each UCB1 seedling rootstock, for example, will differ to some degree in the expression of the P. atlantica genes present in its heritage compared to that of the P. integerrima parent. Greater expression of the P. atlantica frost-tolerance genetics would likely confer greater frost tolerance to the tree.
Avoid budding late-planted rootstocks late
Trying to graft newly planted trees and grow them to the top of the stake in a given season may be a mistake in low elevation areas. If you bud them late, you will have to irrigate the trees late to ensure the buds take. This places trees at increased risk from freezing. It may be better to hold off budding until next year, and reduce irrigation earlier, to decrease tree vigor.
Reduce tree vigor and hypoxia through reduced late season irrigation
As discussed above, years of observations related to WJTD, suggest that trees in “wetter” soils during fall are at greater risk. Whether this is because wetter soils allow the juvenile trees to grow vigorously into the winter without hardening off and placing them at greater risk from a sudden freeze event, or if the roots become hypoxic/anoxic, stressing the tree, and placing it at greater susceptibility to a sudden freeze event, is unknown. Whatever the mechanism, reducing the vigor of the trees by making sure that the root zone is dry prior to the arrival of freezing temperature appears to be an effective, but not a total, preventative of WJTD. On deeper, heavier soils, the last irrigation of juvenile trees should probably occur about the first week of August. If the soils have a high water holding capacity, especially combined with high salinity, it may not be possible to dry them out sufficiently to halt growth by the end of October, but even slowing the growth appears to be beneficial. On shallower, lighter soils, the grower can probably wait longer and may want to reduce irrigation more slowly. Cutting off the water may reduce late-season growth, but data collected in an orchard with deep soils showed no difference in trunk circumference between well-watered trees and those where water was stopped in mid-August after three years of the experiment.
Since pistachio trees, on the existing rootstocks, do not appear to become fully dormant until the end of December, do not irrigate until January, once you shut off the water in August. In saline areas, wait until the new year to begin leaching salt. Current observations suggest the tree is more in danger of frost, than salt. Of course, in a high rainfall year, cutting off irrigation will be less effective, since the orchard may remain wet. Rain, over a large geographical area, however, will increase fog, which will keep nighttime low temperatures higher. Cutting irrigation to fifth and sixth leaf trees is problematic, since drought stress after June will reduce the split, inshell nut percentage appreciably.
If the irrigation system in the orchard is capable of wetting all or most of the ground, running water to keep temperatures above freezing, as is done in citrus, may be a viable alternative. Citrus growers are advised that to run water during or prior to a freeze event, the ground should be firmly packed, free of heavy weed growth/cover crops and the irrigation system should be able to wet more than half the ground area of the orchard. .However, most pistachio growers do not have systems that wet enough of the ground for this to be an effective strategy. Pistachio growers are at a further disadvantage in that the production area for citrus is located in the warmer areas of the SJV, with good cold drainage and not at the bottom of old lake beds where pistachio is grown and where cold air puddles.
The reasons that first and second-leaf trees appear less at risk from WJTD than slightly older trees may be that their root systems are not that well established yet under drip systems, and the tree may be able to be put into drought quicker. The trees, generally, are shorter, where radiant heat from the ground may be more effective at warming them sufficiently to prevent freezing.
Reduce late-season tree vigor through nitrogen management
Do not over apply nitrogen to young trees. Test the soil to see how much nitrogen is present and then add just enough so that little N remains in the soil at the end of the season. Pistachio is an effective scavenger of nitrogen due to its extensive root system. Cut off nitrogen applications in late June or early July.
Reduce hypoxia by improving water infiltration and drainage
Prior to planting, deep rip, plow or chisel soils to breakup subsurface layers that may impede water infiltration and deep drainage. Installation of interceptor trenches for water that my drain from neighboring reservoirs, canals or orchards (either surface of subsurface) or installation of tiles in swampy areas may reduce root hypoxia and improve tree cold tolerance.
Improving infiltration and drainage is difficult on many of the salt-affected soils on the Westside the SJV. The addition of soil amendments such as gypsum or acids in silty, calcareous, and layered soils may not substantially improve water infiltration, but should improve the ability of the plant to absorb calcium in high sodium soils. Calcium has been shown to improve frost tolerance in many crops.
Leaf canopy defoliation
Early defoliation with foliar zinc sulfate sprays or by some other means has been suggested, for many years, as a means of encouraging dormancy. However, scientific research supporting this practice is absent. It does make sense that defoliating the tree should send a message to the tree to slow down. Since freeze events can occur, especially at low elevations at the end of October. Trees, if they are to be chemically defoliated, should be defoliated in mid-October. Defoliation, typically done with zinc sulfate, if it is done at all, should be done in conjunction with soil drying. Trees should not be defoliated too early, as the leaves may regrow, which may reinvigorate the tree, creating a condition that we are trying to avoid. Experimentally defoliating trees with a range of materials has been attempted, but freeze events have not cooperated with the experimenters. Having a dry root zone and a defoliated canopy should take the vigor out of even the most stubborn tree and drought-stressed trees are defoliated much easier than fully irrigated trees.
Use of Tree Wraps?

In a trial, located in a pistachio block on heavy, salty soil west of Lerdo, CA, twenty trees were wrapped with ¾-inch thick, closed cell foam on October 7, 2011 (see picture to left). The wraps extended from the ground up past the graft union. A small temperature recorder was placed inside and outside of three of these wraps approximately12 inches above ground level in early October.
Air temperatures dropped rapidly both inside and outside the wraps in the evening, but low nightly temperatures were approximately 4 ºF colder outside the wraps than inside (see Figure 5 below). Temperatures measured twelve inches above ground level were 4 to 5 ºF colder at their coldest than at four feet above ground level. Note, also, that the temperatures inside the wrap were warmer than those outside the wrap during the warmest part of the day.
The black-coloring of the wraps, and perhaps, proximity to the soil, appeared to increase the daily maximum temperatures, especially inside the wraps, 7 to 12 ºF compared to what the recorder located at four feet above ground level inside of a white, ventilated weather-proof box was measuring (data not shown). Use of white foam wraps would be advised if the grower plans to leave them on through the summer. If the weak point in juvenile pistachio trees is the graft union, a good case could be made for using a wrap like this to help prevent WJTD.

What to do if the trees show WJTD?
Previous experience has shown that the trunks, including the rootstock and scion, of frozen trees, may sunburn in the spring, after the loss of the shading canopy. Whitewash the trunk, especially on the south and west sides of the tree, to prevent sunburn.
Do not try to prune the tree during the winter or early spring. Damaged trees will likely continue to die back during the hotter temperatures of May and June. By late May or June, you should have a good idea if a tree is worth saving.
If the tree has died back to the lower trunk, it is probably best to replace the tree or rebud on a sucker originating from below ground level. Often the trunks are damaged severely, and a stronger tree will likely result if no old scar tissue exists between the producing canopy and the roots.
Old Frost Damage Can Keep Killing Trees

Additional observations related to freeze injury made on farm calls since 2014
In pistachio, as discussed above, most freeze injury is apparent in the spring, shortly after temperatures warm and the trees begin to push. Freeze damage mostly affects juvenile trees. In citrus, we commonly see another type of freeze injury that is slower in developing. In citrus, especially young plants, the tree may be injured near ground level by a severe freeze-event. At the time of the event, the tree shows aerial freeze damage, but appears to regrow normally. In some cases, however, a slow-growing fungus, such as a Fusarium species will start to grow in the freeze-damaged tissue, usually near the crown of the tree. With citrus, after a freeze, growers are advised to prune into green wood if the tree is only partially damaged, to make sure all the blackened, freeze-damaged tissue is removed. Generally, the fungal organisms that invades freeze-damaged wood are the same kind of organism that can invade healthy tissue damaged by a gopher or a disk blade. These organisms grow slowly and the invaded tissue is characterized by a dark discoloration of the wood, although recently into the wood, well below the outer bark and cambium. During farm calls (summer of 2014), pistachio trees were observed with similar symptoms to what we see in citrus, except there was a lot of gumming. All were located in cold areas, and at least one of these orchards had suffered a high percentage of WJTD following a particularly severe freeze event a few years earlier. Typically, if the tree is examined early enough, the scion (i.e. the Kerman or Golden Hills or Peters top) is still alive but gumming, but the rootstock wood is deeply discolored and dead – starting near ground level It can take months and even years for this type of fungus to kill a tree. The trees appear to die when enough of the water-conducting tissues are damaged and the tree cannot keep up with the high late spring transpiration demand.
To find affected trees, look for gumming on the lower trunk of the scion, and then dig around the crown of the tree. The bark will look darkened or dirty in this area. Use a knife or chisel to peel away the bark and scrape the wood beneath. The darkened color of the wood will be obvious. There is no cure for a tree with this kind of damage. The good news is that a healthy replant with an undamaged root system, usually, can be planted in the same hole, with little additional pre-planting preparation. Fusarium species and other soil-borne fungi, grow best in wet environments. To reduce this type of problem, avoid having irrigation water wet or puddle around the crown of the tree.

Citations/References/Further Reading
Baldocchi, D., and E. Waller (2014). Winter fog is decreasing in the fruit-growing region of the Central Valley of California, Geophys. Res. Lett., 41, 3251–3256, https://doi:10.1002/2014GL060018
Charrier, G., J. Ngao, M. Saudreau and T. Ameglio. 2015. Effects of environmental factors and management practices on microclimate, winter physiology, and frost resistance in trees. Front.in Plant Sci. https://doi.org/10.3389/fpls.2015.00259
Chartzoulakis, K., M. Loupassaki, M. Bertaki, and I. Androulakis. 2002. Effects of NaCl salinity on growth, ion content and CO2 assimilation rate of six olive cultivars. Scientia Horticulturae 96: 235–247.
Chartzoulakis, K, G Psarras, S. Vemmos, M. Loupassaki, 2006. Response of two olive cultivars to salt stress and potassium supplement. Journal of Plant Nutrition 29: 2063–2078.
Ferguson, L., J. Poss, S. Grattan, C. Grieve, D. Wang, C. Wilson, T. Donovan and C. Chao. 2002. Pistachio rootstocks influence scion growth and ion relations under salinity and boron stress. J. Amer. Soc. Hort. Sci. 127(2):194-199.
Heil K and U. Schmidhalter. 2017. The Application of EM38: Determination of Soil Parameters, Selection of Soil Sampling Points and Use in Agriculture and Archaeology. Sensors (Basel). 2017;17 (11):2540. Published 2017 Nov 4. https://doi:10.3390/s17112540
Jan, N., M. ul-Hussain and K. Andrabi. 2009. Cold resistance in plants: A mystery unresolved. Electronic Journal of Biotechnology, Vol. 12 No. 3, Issue of July 15, 2009. https://doi:10.2225/vol12-issue3-fulltext-3
Kallsen, C.E., 2012. Reducing Juvenile Pistachio Tree Damage. https://www.acpistachios.org/Industry/research/7-2012.pdf
Li, P. H., and C.J. Weiser. 1971. Increasing cold resistance of stem sections of Cornus stolonifera by artificial dehydration: a preliminary report. Cryobiology 8, 108–111. https://doi.org/10.1016/0011-2240(71)90111-8
Loupassaki, M.H., K.S. Chartzoulakis, N.B. Digalaki, and I.I. Androulakis. 2002. Effect of salt stress on concentration of nitrogen, phosphorous, potassium, calcium, magnesium, and sodium in leaves, shoots, and roots of six olive cultivars. J. of Plant Nutrition 25:2457-2482.
Melgar, J.C., M. Benlloch, and R. Fernandez-Escobar. 2006. Calcium increases sodium exclusion in olive plants. Scientia Horticulturae 109:303-305.
Morgan, P.W. and M.C. Drew. 2006. Ethylene and plant responses to stress. Physiologia Plantarum. https://doi.org/10.1111/j.1399-3054.1997.tb03068.x
Palta, J.P. 1996. Role of calcium in plant responses to stresses: Linking basic research to the solution of practical problems. HortScience 31:51–57.
Palta, J.P., and P.H. Li. 1980. Alterations in membrane transport properties by freezing injury in herbaceous plants. Evidence against rupture theory. Physiologia Plantarum 50:169–175.
Pellet, N.E., and D.B. White. 1969. Effects of soil nitrogen and soil moisture levels on cold acclimation of container grown Juniperus chinensis ‘Hetsi’. J. Amer. Soc. Hort.Sci. 94:457–459.
Percival, G.C., C. Boyle and L. Baird. 1999. The influence of calcium supplementation on the freezing tolerance of woody plants. J. of Arboriculture 25(6): 285-291
Percival, G.C. and S. Barnes. 2008. Calcium-induced freezing and salinity tolerance in evergreen oak and apple cv. ‘Golden Crown’. Arboriculture & Urban Forestry 34(3):191–199.
Salazar, C., C. Hernandez and M.T. Pinol. 2015. Plant water stress: Associations between ethylene and abscisic acid response. Chilean Jour. of Agric. Res. (Suppl. 1) pgs. 71-79. August, 2015 79
Sperling, O, F. Secchi, J. Godfrey, and M. A. Zwieniecki, (2016). Acclimation of Pistacia integerrima trees to frost in semi-arid environments depends on autumn’s drought. Planta. 245. 10.1007/s00425-016-2629-9.